 |
Slovenská arachnologická spoločnosť |
|
|
|
|
|
|
|
|
|
Pavúkovce (Arachnida) sú nápadnou a početnou živočíšnou skupinou. Obývajú prakticky každý suchozemský
habitat, s niekoľkými zástupcami v sladkej vode a v mori. Všeobecne sú to
dravce, ale niektoré z nich sú fytofágy a iné zasa obligátne parazity
živočíchov alebo rastlín. Veľkosť tela dospelých zvierat varíruje od 250 μm pri roztočoch po veľké
diskovité „tarantuly”.
Medzi pavúkovce patria niektoré z najjedovatejších živočíchov na Zemi – niektoré pavúky a škorpióny sú schopné za istých okolností zabiť dospelého človeka – a mnohé vyvolávajú
strach a odpor celých ľudských kultúr. Niektoré roztoče a kliešte sú vektormi chorôb, ktoré spôsobujú veľké problémy
mnohým ľuďom. Väčšinou sú ale pavúkovce pre človeka neškodné a nebiológmi sú videné iba vzácne.
Arachnida obsahujú 11 recentných skupín (ak roztoče vnímame na úrovni radu). Pavúkovce
sú starobylými živočíchmi. Najmenej tri recentné skupiny sa objavujú vo fosílnych záznamoch v silúre a devóne a väčšinu zostávajúcich
skupín poznáme z
karbónu.

|
Acari |
 |
54 781
recentných druhov |
|
opistozomálna segmentácia nezreteľná alebo chýba; rozdelenie tela na prozómu a opistozómu nie je zreteľné |

|
|
Amblypygi |
 |
161
recentných druhov |
|
opistozomálna segmentácia zreteľná, telo sploštené, opistozóma bez predĺženia, palpy raptoriálne |

|
|
Araneae |
 |
47 743
|
|
opistozomálna segmentácia nezreteľná alebo chýba; rozdelenie tela na prozómu a opistozómu je zreteľné |

|
|
Opiliones |
 |
6 653
|
|
opistozomálna segmentácia zreteľná, opistozóma bez predĺženia, palpy jednoduché; rozdelenie tela na prozómu a opistozómu nie je zreteľné |

|
|
Palpigradi |
 |
82
recentných druhov |
|
opistozomálna segmentácia zreteľná, opistozóma zakončená segmentovaným flagellom, palpy jednoduché |
|
|
Pseudoscorpiones |
 |
3 454
|
|
opistozomálna segmentácia zreteľná, opistozóma bez predĺženia, palpy klieštikovité |
|
|
Ricinulei |
 |
58
recentných druhov |
|
opistozomálna segmentácia zreteľná, prosoma s oddeleným článkom (cucullus) pred karapaxom, oválna opistozóma bez predĺženia, palpy klieštikovité |
|
|
Schizomida |
 |
260
recentných druhov |
|
opistozomálna segmentácia zreteľná, pedicel prítomný, opistozóma s krátkym flagellom, palpy robusné a jednoduché, prvý pár nôh slúži ako senzorický orgán |
|
|
Scorpiones |
 |
2 323
|
|
opistozomálna segmentácia zreteľná, opistozóma predĺžená (metasoma) so žihadlom na konci, palpy klieštikovité |

|
|
Solifugae |
 |
1 113
|
|
opistozomálna segmentácia zreteľná, opistozóma bez predĺženia, palpy jednoduché, chelicery mohutné |
|
|
Thelyphonida |
 |
110
recentných druhov |
|
opistozomálna segmentácia zreteľná, opistozóma predĺžená segmentovaným flagellom; palpy raptoriálne |
|

Fosílne pavúkovce
|
Trigonotarbida |
 |
68
fosílnych druhov |
|
na prvý pohľad pripomínajú pavúky, ale na chrbtovej strane opistozómy majú tergity, rozdelené ešte na mediánne a laterálne štítky |
|
|
stavba tela pavúkovcov |
|
Hlava pavúkovcov sa nikdy osobitne nevyvíja a nie je nikdy oddelená od hrudných článkov. Dokonca aj „mozog“ (v skutočnosti nadhltanový a pohltanový nervový uzol) je uložený skôr niekde v oblasti prvých párov nôh. Preto je vhodnejšie nazývať oddiely tela pavúkovcov prozóma a opistozóma. Predná časť prozóma [pro] je zhora krytá pancierom (karapax alebo epistóm [epi]) a nesie 6 párov končatín. Na karapaxe sa môžu nachádzať mediánne a bočné oči. Zadná časť opistozóma je pôvodne zložená z 12 článkov, ale väčšina skupín pavúkovcov má redukovaný počet článkov. Opistozóma môže byť ešte rozdelená na širšiu mezozómu [mes] a užšiu metazómu [met]. Na mezozóme fosílnych pavúkovcov sa nachádzali žiabrové prívesky [zp]. Spojenie opistozómy s prozómou zabezpečuje rôzne modifikovaný siedmy článok, napríklad vo forme stopky (pedicel). |
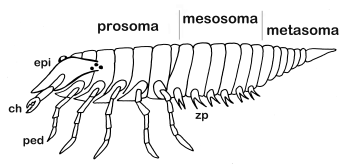 stavba tela hypotetického predka pavúkovcov
stavba tela hypotetického predka pavúkovcov© Martin Mrva (in: Matis et al. 2003) |
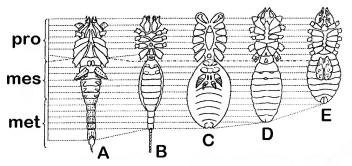 schéma stavby tela pavúkovcov: A – Scorpiones, B –
Thelyphonida, C – Araneae,
schéma stavby tela pavúkovcov: A – Scorpiones, B –
Thelyphonida, C – Araneae, D – Solifugae, E – Opiliones. Prerušovanou čiarou je naznačený 7. článok tela. © Martin Mrva (in: Matis et al. 2003) |
Prvým párom končatín sú chelicery (klepietka) [ch], ktoré slúžia na chytanie či trhanie potravy a sú obyčajne klepietkovitého tvaru (výnimkou sú napríklad pavúky či mnohé roztoče). Bývajú dvoj- alebo trojčlánkové (pôvodnejší typ, Palpigradi, Opiliones, Scorpiones). Chelicery šťúrikov obsahujú snovaciu žľazu, chelicery pavúkov zasa jedovú žľazu. Druhým párom končatín sú pedipalpy (hmatadlá, palpy) [ped], ktoré majú zmyslovú funkciu (obdoba tykadiel hmyzu) a podobajú sa nohám. Vždy sú zložené zo 6 článkov. Pri mnohých pavúkovcoch sú nápadne pozmenené na chytanie potravy (veľmi nápadne premenené na klepetá majú škorpióny a šťúriky, na chytavé štruktúry amblypygi, uropygi či niektoré roztoče). Pedipalpy môžu mať aj rôzne špecializované funkcie – samce pavúkov ich používajú ako kopulačný orgán a solifúgy majú pedipalpy ukončené prísavkami. |
|
Za klepietkami a hmatadlami nasledujú na prozóme štyri páry kráčavých nôh, ktoré slúžia zväčša na pohyb. Prvý pár nôh niekedy preberá hmatovú funkciu (amblypygi, tartaridy, solifúgy a niektoré roztoče) a nie je používaný na pohyb. Nohy majú 6 článkov – coxa (panvička), trochanter (predpanvička), femur (stehno), genu (koleno, niekedy ako patella), tibia (holeň) a tarsus (chodidlo). Niektoré články môžu byť ešte druhotne rozdelené. Panvičky môžu mať výrastky (nazývané maxillae alebo gnathobasae), ktoré slúžia na mechanické spracovanie potravy (pri niektorých koscoch sú to všetky panvičky, pri škorpiónoch iba panvičky prvých dvoch párov nôh a pri pavúkoch sa nohy vôbec nepodieľajú na mechanickom spracovaní potravy). Ak porovnáme nohu pavúkovcov a hmyzu, pavúkovce majú o jeden článok viac (genu). Za deriváty brušných končatín považujeme pohlavné uzávery (operculum) škorpiónov a snovacie bradavky pavúkov. Medzi deriváty pokožky zaraďujeme jedové žľazy (Araneae, Pseudoscorpiones, Scorpiones) a snovacie žľazy (Araneae, Pseudoscorpiones, Acariformes). Samce niektorých pavúkovcov využívajú ako kopulačný orgán končatiny (pedipalpy pri Araneae, tretí pár nôh pri Ricinulei). Skoro všetky pavúkovce majú schopnosť regenerácie končatín, ktoré stratia v nymfálnom štádiu. |
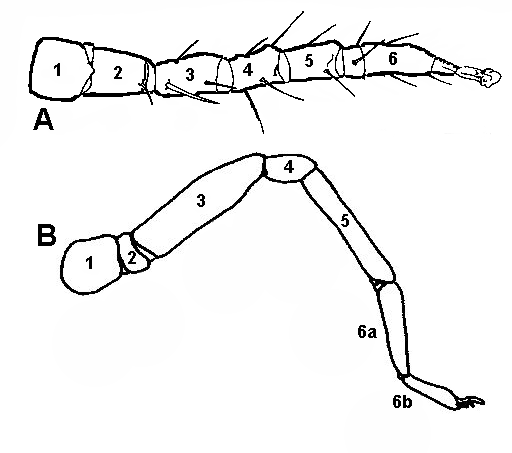 schémy stavby nohy pavúkovcov: A – Acari, B – Araneae
schémy stavby nohy pavúkovcov: A – Acari, B – Araneae (1 – coxa, 2 – trochanter, 3 – femur, 4 – genu, 5 – tibia, 6 – tarsus, 6a – metatarsus, 6b – acrotarsus) |
Tráviaca sústava je podobná ako pri ostatných článkonožcoch. Príjem potravy
je externý. Zo stredného čreva (mesenteron) vybiehajú početné výbežky (diverticula), ktoré slúžia čiastočne
aj ako zásobný orgán a vďaka ktorému dokážu pavúkovce vydržať dlhé obdobie bez potravy.
Vylučovacia sústava využíva rôzne spôsoby vylučovania. Najlepšie je vyvinutý systém
koxálnych žliaz.
Malpigiho trubice pavúkovcov nie sú homologické s hmyzom (pri Arachnida sú hypodermálneho pôvodu).
Dýchajú pľúcnymi vakmi alebo vzdušnicami (tracheálny systém má ale inú štruktúru ako má napríklad hmyz).
Tracheálny systém majú Pseudoscorpiones, Solifugae, Opiliones, Ricinulei a Acari.
Pľúcne vaky (1, 2 alebo 4 páry) majú
Scorpiones, Uropygi a Amblypygi. Väčšina Araneae má pľúcne vaky aj tracheálny systém. Drobné Palpigradi a
niektoré Acari nemajú dýchacie orgány a dýchajú povrchom tela. Nervová sústava je podobná ako pri ostatných článkonožcoch.
Pohlavná sústava pavúkovcov vykazuje nezvyčajnú variabilitu.
Sú oddeleného pohlavia, častý je
pohlavný dimorfizmus.
Samce Opilionida a Acariformes majú penis. Častý je prenos spermií
spermatofórmi. Samice majú často vyvinutú starostlivosť o potomstvo (nosenie prvých
instarov na chrbte samicami škorpiónov a niektorých pavúkov, vak (marsupium) na brušnej strane samíc štúrikov).
Vývin pavúkovcov obsahuje vajíčko, larvu, nymfálne a dospelé štádiá. Všeobecne majú pavúkovce tri nymfálne štádiá: protonymfa – deutonymfa – tritonymfa,
ktoré sú oddelené zvliekaniami. Poznáme ale skupiny pavúkovcov, kde je počet nymfálnych štádií redukovaný, či na druhej strane pri iných skupinách môže byť
až dvanásť zvliekaní (pri pavúkoch je známych dokonca až 22 zvliekaní, pričom počet zvliekaní aj v rámci druhu môže varírovať).
Pavúkovce žijú asi rok až 25 rokov (niektoré mygalomorfné pavúky).
Zachvatkin (1952) navrhol rozdelenie pavúkovcov na tri skupiny podľa prítomnosti opticky aktívneho chitínu v povrchových štruktúrach tela. Opticky aktívny chitín znamená, že je schopný otáčať rovinu polarizovaného svetla. Holoactinochitinosi majú opticky aktívny chitín ako v ochlpení, tak aj v telovom pokryve (rad Scorpiones). Prítomnosť opticky aktívneho chitínu v kutikule ale nespôsobuje známe svetielkovanie škorpiónov v UV svetle (za to môžu alkaloid β-karbolín a fenylpropanoid 7-hydroxy-4-metylkumarín). Actinochaeta (solifúgny typ) majú opticky aktívny chitín iba v chĺpkoch na tele (rady Solifugae, Palpigradi, Schizomida, Thelyphonida, Pseudoscorpionida, roztoče radu/nadradu Acariformes). Actinoderma (arachnoidný typ) vôbec nemajú opticky aktívny chitín (rady Araneae, Opiliones, Amblypygi, Ricinulei, roztoče radu/nadradu Parasitiformes). |
 škorpión v ultrafialovom svetle
škorpión v ultrafialovom svetle |
Pavúkovce prejavujú fylogenetickú tendenciu k splývaniu článkov jednotlivých častí tela, takže dochádza ku koncentrácii nervovej sústavy, ktorá je mimoriadne výkonná už vzhľadom na prevládajúci dravý spôsob života. Prejavuje sa to aj na formovaní mnohých inštinktov, ktoré sú u pavúkovcov veľmi nápadným javom.
viac informácií:
![]()
![]()
|
klasifikácia a fylogenéza pavúkovcov |
Pavúkovce (Arachnida) sú jednou z piatich hlavných skupín podkmeňa klepietkavcov (Chelicerata). Prehľad tried klepietkavcov v slovenčine nájdu záujemcovia na blogu vivere est viventibus esse. Pavúkovce sú suchozemské živočíchy, tradične rôznymi kultúrami považované za nebezpečné a tajomstvami opradené živočíchy. Dnes sa väčšina morfológov, paleontológov a molekulárnych taxonómov zhoduje, že článkonožce (Arthropoda) sú monofyletický taxón a klepietkavce sú nesporne ich osobitnou vývinovou vetvou. V minulosti sa predpokladalo, že trilobity (Trilobitomorpha) sú vetvou vybiehajúcou z hlavnej línie klepietkavcov (Weygoldt 1998). Dnes prevláda názor, že Trilobitomorpha a Chelicerata sú zrejme príbuzné evolučné línie, ale žiadna z nich nie je predkom tej druhej. Geologické a morfologické dôkazy naznačujú, že prvé šťúry boli prinajmenšom čiastočne vodnými živočíchmi a prechod pavúkovcov na suchú zem sa v evolúcii uskutočnil viac ako jeden krát nezávisle od seba (škorpióny a jedna alebo aj viac ďalších línií pavúkovcov). Prvá adaptívna radiácia pavúkovcov prebehla veľmi skoro, pretože všetky súčasné rady pavúkovcov sa vyskytovali už v karbóne (teda pred 358–298 miliónmi rokov).
Dnes existuje široká zhoda odbornej verejnosti pri definícii recentných radov pavúkovcov. Desať radov je prijímaných bez výhrad, problematickým taxónom zostávajú iba roztoče. Existujú tri názory – Acari na úrovni radu, Acari na úrovni podtriedy s dvoma radmi Parasitiformes a Acariformes (hlavne v prácach evolučných biológov) alebo so šiestimi radmi Opilioacarida, Holothyrida, Ixodida, Mesostigmata, Trombidiformes a Sarcoptiformes (súčasné práce akarológov používajú koncept 6 radov roztočov). Štyri rady pavúkovcov sú známe iba z fosílneho záznamu – † Haptopoda (1 druh), † Phalangiotarbida (31 druhov), † Trigonotarbida (65 druhov) a † Uraraneida (3 druhy). Grimaldi & Engel (2005) spomínajú ešte tri fosílne taxóny – † Sidneyiida, † Emeraldellida a † Sanctacarida. Zhang (2011) ich zaradenie do súčasného systému považuje za nemožné kvôli neúplnému fosílnemu záznamu. |
 Kreischeria (Trigonotarbida)
Kreischeria (Trigonotarbida) Naturhistorisches Museum vo Viedni |

|
Na stránke Medzinárodnej arachnologickej spoločnosti sa nachádza
zoznam fosílnych klepietkavcov (Dunlop et al. 2015).
Fosílie sú uvedené a zaradené v rámci moderných, v súčasnosti bežne uznávaných systémov (Norman Platnick, Victor Fet, Mark Harvey, Adriano Kury, manuál akarológie).
Vyhynuté čeľade sú zaradené na pozície, kde podľa autorov najlepšie odzrkadľujú ich príbuzenské vzťahy.
Zoznam obsahuje aj citácie všetkých originálnych opisov, a kde to je možné je pridaná aj relevantná taxonomická literatúra.
|
Acaromorpha = Ricinulei + Acari |
 |
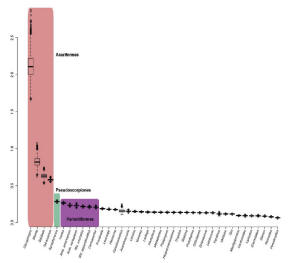 Bayeziánsky test relatívných rýchlostí. (podľa Sharma et al. 2014)
Bayeziánsky test relatívných rýchlostí. (podľa Sharma et al. 2014) |
Monofyletický pôvod pavúkovcov je podporený najmenej 11 synapomorfickými znakmi (Coddington et al. 2004). Vzhľadom na to, že paleontologický materál pavúkovcov je relatívne obsiahly – Dunlop (2010) a Garwood & Dunlop (2014) uvádzajú asi 2 200 valídnych fosílnych druhov – existuje množstvo fylogenetických hypotéz. Prvé vznikali na základe morfologických znakov, neskôr na základe ribozomálnych alebo jadrových génov. Dnešné pohľady na fylogenézu recentných pavúkovcov považujú za najprimitívnejšie skupiny Ricinulei a Scorpiones. Pôvod Palpigradi je nejasný a zostáva naďalej záhadou. Samostatnú vetvu tvorí skupina Tetrapulmonata (teda Araneae a skupina Pedipalpi). Fylogenetické postavenie ostatných radov je stále predmetom diskusií. Dabert et al. (2010) napríklad molekulárnymi dátami určili ako sesterské skupiny Acariformes–Solifugae a Parasitiformes–Pseudoscorpionida. Sharma et al. (2014) uvádzajú, že najodvodenejšie rady pavúkovcov sú zároveň skupinami s najväčším množstvom mutácií, kde je evolúcia zrýchlená (na základe Bayeziánskeho testu relatívnych rýchlostí 500 pomaly sa vyvíjajúcich génov). Najväčší počet mutácií je v rade Acariformes (obrázok vľavo, červený stĺpec), nasledujú Pseudoscorpiones (zelený stĺpec) a Parasitiformes (fialový stĺpec). Tento stav sa odráža aj v biodiverzite týchto radov, z veľkej časti ešte stále nepoznanej. Schematický a idealizovaný fylogenetický strom súčasných pavúkovcov odzkadľujúci dnešné názory: |
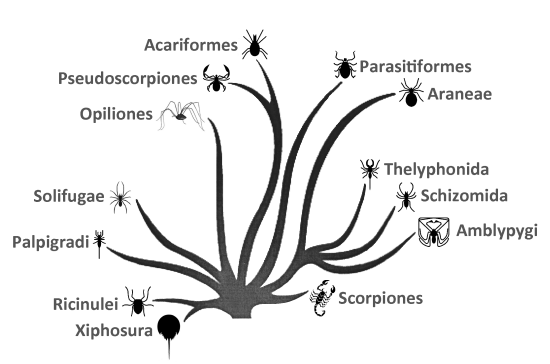
|
diverzita pavúkovcov |
Všetky rady pavúkovcov je možné na základe počtu opísaných druhov rozdeliť do troch skupín (Harvey 2002). Megadiverzifikované rady – pavúky a rady roztočov – tvoria spoločne skoro 88% opísaných druhov pavúkovcov. Pavúky majú dnes 47 743 opísaných druhov (World Spider Catalog 2018) a roztoče 54 781 druhov (rovnako sú megadiverzifikované aj oba rady roztočov – Parasitiformes s 13 005 druhmi a Acariformes s 41 776 druhmi; prípadne podľa dnes prevládajúcej klasifikácie rady Mesostigmata s 12 017 druhmi, Trombidiformes s 25 146 druhmi a Sarcoptiformes so 16 630 druhmi) (Walter & Proctor 2013). Mezodiverzifikované rady – Opiliones, Pseudoscorpiones, Scorpiones a Solifugae – obsahujú každý viac ako 1 000 pomenovaných druhov, ale nedosahujú a ani nedosiahnu úroveň diverzity pavúkov a roztočov. Kosce sú z nich druhovo najbohatšie s 6 484 opísanými druhmi, nasledujú šťúriky s 3 454 opísanými druhmi, škorpióny s 2 323 druhmi a solifúgy s 1 113 opísanými druhmi (Zhang 2011, Rein 2016). Ak budeme pokladať kliešte (Ixodida) za rad, tak s 918 druhmi ich musíme tiež zaradiť do tejto skupiny.
Mikrodiverzifikované rady – Schizomida, Amblypygi, Uropygi, Palpigradi
a Ricinulei – obsahujú niektoré z najviac geograficky ohraničených taxónov článkonožcov
vôbec (Harvey 2002). S výnimkou tartaríd žiaden z nich neobsahuje ani len 200 opísaných druhov.
Ak chápeme roztoče zo skupín Opilioacarida a Holothyrida ako rady, tak
patria tiež medzi mikrodiverzifikované rady. |
 Ernst Haeckel: Kunstformen der Natur,
Plate 66 (1904)
Ernst Haeckel: Kunstformen der Natur,
Plate 66 (1904) |
|
základné informácie o pavúkovcoch |
Základnú monografiu o pavúkovcoch pre celú generáciu arachnológov zostavil Theodore H. Savory. Vyšla v roku 1964, druhé doplnené vydanie vyšlo v roku 1977 (Savory 1964, 1977). Janet Beccaloni o štyridsať rokov neskôr vtedajšie vedomosti o svete pavúkovcov interpretovala prístupným jazykom pre širšiu verejnosť (Beccaloni 2009). Kniha je ilustrovaná zaujímavými a často bizardnými faktami a viac ako 200 fotografiami a kresbami. Aktuálne základné informácie o pavúkovcoch v slovenčine sa nachádzajú hlavne vo vysokoškolských skriptách jednotlivých prírodovedeckých fakúlt – v Bratislave je to Matis et al. (2003), v Košiciach Panigaj & Ľuptáčik (2015), v Banskej Bystrici Franc (2005). Rozsiahly a hodnotný text venovaný pavúkovcom obsahuje poľská učebnica bezstavovcov, ktorú zostavil známy akarológ Czesław Błaszak (2011). Petar Beron ponúkol zaujímavý pohľad na pavúkovce tým, že skombinoval poznatky o rozšírení všetkých skupín pavúkovcov s geografickými údajmi (Beron 2018).




Beccaloni, J. 2009. Arachnids. The Natural History Museum, London, 320 pp.
Beron, P. 2018. Zoogeography of Arachnida. Monographiae Biologicae 94, Springer International Publishing, 988 pp.
Błaszak, C. (ed) 2011. Zoologia. Stawonogi. Tom 2, część 1. Szczękoczułkopodobne, skorupiaki. Wydawnictwo Naukowe PWN, Warszawa, 407 pp.
Coddington, J.A., Giribet, G., Harvey, M.S., Prendini, L., Walter, D.E. 2004. Arachnida. In: Cracraft, J., Donoghue, M.J. (eds) Assembling the Tree of Life. Oxford University Press, pp. 296–318.
Dabert, M., Witaliński, W., Kazmierski, A., Olszanowski, Z., Dabert, J. 2010. Molecular phylogeny of acariform mites (Acari, Arachnida): strong conflict between phylogenetic signal and long-branch attraction artifacts. Molecular Phylogenetics and Evolution 56 (1): 222–241.
Dunlop, J.A. 2010. Geological history and phylogeny of Chelicerata. Arthropod Structure & Development 39 (2–3): 124-142.
Dunlop, J.A., Penney, D., Jekel, D. 2015. A summary list of fossil spiders and their relatives. In World Spider Catalog. Natural History Museum Bern, online at http://wsc.nmbe.ch, version 16.0, accessed on 15. September 2018.
Franc, V. 2005. Systém a fylogenéza živočíchov – bezchordáty (Doplnená prepracovaná verzia, Vol. 2). Fakulta prírodných vied, Univerzita Mateja Bela v Banskej Bystrici, 149 pp.
Garwood, R.J., Dunlop, J. 2014. Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders. PeerJ 2: e641, pp. 1–33.
Grimaldi, D., Engel, M.S. 2005. Evolution of the Insects. Cambridge University Press, Cambridge, New York.
Harvey, M.S. 2002. The neglected cousins: what do we know about the smaller arachnid orders? The Journal of Arachnology 30: 357–372.
Harvey, M.S. 2013a. Palpigrades of the World, version 1.0. Western Australian Museum, Perth, online at http://www.museum.wa.gov.au/catalogues/palpigrades, accessed on 15. October 2016.
Harvey, M.S. 2013b. Pseudoscorpions of the World, version 3.0. Western Australian Museum, Perth, online at http://museum.wa.gov.au/catalogues/pseudoscorpions, accessed on 15. October 2016.
Harvey, M.S. 2013c. Ricinuleids of the World, version 1.0. Western Australian Museum, Perth, online at http://www.museum.wa.gov.au/catalogues/ricinuleids, accessed on 15. October 2016.
Harvey, M.S. 2013d. Schizomids of the World, version 1.0. Western Australian Museum, Perth, online at http://www.museum.wa.gov.au/catalogues/schizomids, accessed on 15. October 2016.
Harvey, M.S. 2013e. Solifuges of the World, version 1.0. Western Australian Museum, Perth, online at http://www.museum.wa.gov.au/catalogues/solifuges, accessed on 15. October 2016.
Harvey, M.S. 2013f. Whip scorpions of the World, version 1.0. Western Australian Museum, Perth, online at http://www.museum.wa.gov.au/catalogues/whip-scorpions, accessed on 15. October 2016.
Harvey, M.S. 2013g. Whip spiders of the World, version 1.0. Western Australian Museum, Perth, online at http://www.museum.wa.gov.au/catalogues/whip-spiders, accessed on 15. October 2016.
Kury, A.B. 2002. Checklist of valid genera of Opiliones of the World. Museu Nacional/UFRJ website, online at http://www.museunacional.ufrj.br/mndi/Aracnologia/checklaniator.htm, accessed on 5. April 2017.
Kury, A.B. 2018. Classification of Opiliones. Museu Nacional/UFRJ website. Online at: http://www.museunacional.ufrj.br/mndi/Aracnologia/opiliones.html, accessed on 10. November 2018.
Matis, D., Krumpál, M., Beláková, A., Fedor, P. 2003. Zoológia bezchordátov 2 diel. Tentaculata, Pogonophora, Onychophora, Arthropoda, Hemichordata, Echinodermata. Vysokoškolské učebné texty pre prírodovedecké fakulty, Faunima, Bratislava, 168 pp.
Panigaj, Ľ., Ľuptáčik, P. 2015. Zoológia bezchordátov (náčrt systému a fylogenézy). Vysokoškolský učebný text pre Prírodovedeckú fakultu UPJŠ v Košiciach, vydavateľ Univerzita Pavla Jozefa Šafárika v Košiciach, 128 pp.
Rein, J.O. 2016. The Scorpion Files. NTNU, online at https://www.ntnu.no/ub/scorpion-files/, accessed on 5. April 2017.
Savory, T.H. 1964. Arachnida. Academic Press Inc., 291 pp.
Savory, T.H. 1977. Arachnida. Second Edition. Academic Press Inc., 348 pp.
Schmidt, K.-H. 2010. AcariBase – database of mites with more than 58.000 names, online at http://www.miteresearch.org/index1.html, accessed on 17. October 2016.
Sharma, P.P., Kaluziak, S.T., Pérez-Porro, A.R., González, V.L., Hormiga, G., Wheeler, W.C., Giribet, G. 2014. Phylogenomic Interrogation of Arachnida Reveals Systemic Conflicts in Phylogenetic Signal. Molecular Biology and Evolution 31 (11): 2963–2984.
Walter, D.E., Proctor, H.C. 1999. Mites: ecology, evolution and behaviour. CABI Publishing, Wallingford, 322 pp.
Walter, D.E., Proctor, H.C. 2013. Mites: ecology, evolution and behaviour – life at a microscale. Second Edition. Springer, The Netherlands, 494 pp.
Weygoldt, P. 1998. Evolution and systematics of the Chelicerata. Experimental & Applied Acarology 22 (2): 63–79.
Word Spider Catalog 2018. World Spider Catalog. Natural History Museum Bern, online at http://wsc.nmbe.ch, version 19.5, accessed on 27. July 2018.
Zachvatkin, A.A. 1952. Razdeleniye kleshchey (Acarina) na otriady i ikh polozheniye v sisteme Chelicerata. Parasitologicheski sbornik Zoologicheskogo Instituta AN SSSR 14: 5–46.
Zhang, Z.-Q. 2011. Phylum Arthropoda von Siebold, 1848 In: Zhang, Z.-Q. (Ed.) Animal biodiversity:
An outline of higher-level classification and survey of taxonomic richness. Zootaxa 3148: 7–12.
![]()
|
|
| © 2016 SARAS, o.z. |
| Posledná aktualizácia: 30. marec 2019 |
|
|